| Dinosaurs
(excluding Aves) | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
 Mounted skeletons of Tyrannosaurus (left) and Apatosaurus (right) at the AMNH.
| ||||||||||||
| Scientific classification | ||||||||||||
| ||||||||||||
|
Dinosaurs are an extinct, diverse, largely terrestrial group of vertebrate animals of the Sauropsid orders Saurischia (lizard-hipped) and Ornithischia (bird-hipped) and were the dominant land reptiles for over 160 million years, during the Mesozoic era. Dinosaurs first appeared approximately 230 million years ago (mya) during the Triassic and disappeared at the end of the Cretaceous period, approximately 65 mya, when a catastrophic extinction event ended their dominance on land. The Mesozoic is often called the "Age of Dinosaurs" after the dominant fauna of the era. Modern birds are generally considered to be the direct descendants of theropod dinosaurs.
The finding of fossils of the "terrible lizards" presented a conundrum for young earth creationists, who maintain that the earth is thousands of years old rather than millions of years old. Initially, a different kind of religious/philosophical problem, related to extinction, was presented to those who professed Divine Creation of all of nature’s creatures: If God is perfect, and He made a perfect creation, why would He allow any of His created organisms to become extinct? However, for most present-day religious adherents, the finding of fossils of dinosaurs from many millions of years ago is not problematic to the view of a Creator God, since it is recognized that all development passes through stages involving time, such as the life cycle of a butterfly (egg, larva, pupa, and adult) or the history of life on Earth (Precambrian, Paleozoic, Mesozoic, Cenozoic, etc.) Rather, earlier stages provide a foundation for future stages. The relatively sudden demise of dinosaurs, after a long period of dominance, provided the foundation for events such as the rise of mammals and birds.
Since the first dinosaur fossils were recognized in the nineteenth century, mounted dinosaur skeletons have become major attractions at museums around the world. Dinosaurs have become a part of world culture and remain consistently popular among children and adults alike. They have been featured in best-selling books and films such as Jurassic Park, and new discoveries are regularly covered by the media.
The term dinosaur is sometimes used informally to describe other prehistoric reptiles, such as the pelycosaur Dimetrodon, the winged pterosaurs, and the aquatic ichthyosaurs, plesiosaurs, and mosasaurs, although technically none of these were dinosaurs.
| Mesozoic era (251 - 65 mya) | ||
|---|---|---|
| Triassic | Jurassic | Cretaceous |
What is a dinosaur?
Definition

The taxon Dinosauria was formally named by the English palaeontologist Richard Owen in 1842 as "a distinct tribe or sub-order of Saurian Reptiles" (Owen 1842). The term is derived from the Greek words δεινός (deinos meaning "terrible", "fearsome," or "formidable") and σαύρα (saura meaning "lizard" or "reptile"). Owen chose the name to express his awe at the size and majesty of the extinct animals (Farlow and Brett-Surman 1997), not out of fear or trepidation at their size and often-formidable arsenal of teeth and claws. Owen was describing this group based on three large fossils found in England: Megalosaurus, Iguanodon, and Hylaeosaurus.
Dinosaurs were an extremely varied group of animals. According to a 2006 study, 527 dinosaur genera have been identified with certainty so far, and 1,844 genera are believed to have existed (Wang and Dodson 2006, Fountain 2006). Some were herbivorous, others carnivorous. Some dinosaurs were bipeds (moving on two legs), some were quadrupeds (moving on four legs), and others, such as Ammosaurus and Iguanodon, are considered to be able to walk just as easily on two or four legs. Regardless of body type, nearly all known dinosaurs were well-adapted for a predominantly terrestrial—that is, land-bound—rather than aquatic or aerial, habitat.
Distinguishing features of dinosaurs
While recent discoveries have made it more difficult to present a universally agreed-upon list of dinosaurs' distinguishing features, nearly all dinosaurs discovered so far share certain modifications to the ancestral archosaurian skeleton. Archosaurs are a group of diapsid reptiles represented today by birds and crocodiles. This group also included the dinosaurs. Diapsids ("two arches") are a group of tetrapod animals that appeared in the fossil record about 300 million years ago during the late Carboniferous period. Diapsids possess a pair of holes in their skulls behind the eyes, along with a second pair located higher on the skull.
Although some later groups of dinosaurs featured further modified versions of these common archosaurian traits, they are considered typical across Dinosauria; the earliest dinosaurs had them and passed them on to all their descendants. Such common structures across a taxonomic group are called synapomorphies.
Dinosaur synapomorphies include: an elongated crest on the humerus (upper arm bone) for deltopectoral muscles; a shelf on the back-rear of the ilium (main hip bone); a tibia (shin) with its lower edge expanded side to side and a flange pointing to the rear and outside; and an ascending projection on the astragalus (one of the ankle bones) which locks it to the tibia (Benton 2004a).

A variety of other skeletal features were shared by many dinosaurs. However, because they were either common to other groups of archosaurs or were not present in all early dinosaurs, these features are not considered to be synapomorphies. Such shared features include a diapsid skull bearing two pairs of holes in the temporal region, and holes in the snout and lower jaw—two characteristics shared by other archosaurs. Other characteristics include loss of the skull's postfrontal bone; a long neck incorporating an S-shaped curve (Holtz 2000); an elongated scapula, or shoulder blade; forelimbs shorter and lighter than hind limbs, coupled to asymmetrical hands; a sacrum composed of three or more fused vertebrae; and an acetabulum, or hip socket, with a hole at the center of its inside surface (Langer et al. 1999).
The open, or "perforate," hip joint of dinosaurs had significant implications for dinosaur movement and behavior. Most notably, it allowed dinosaur hind limbs to be underslung, or situated directly beneath the animals' bodies; this, in turn, allowed dinosaurs to stand erect in a manner similar to modern mammals, but distinct from most other reptiles, whose limbs sprawl out to either side (Benton 2004b). Vertical limb configuration also enabled dinosaurs to breathe easily while moving, which likely permitted stamina and activity levels that surpassed those of sprawling reptiles.
Taxonomic definition
Under phylogenetic taxonomy (classification centered on evolutionary relatedness), dinosaurs are defined as descendants of the most recent common ancestor of Triceratops and modern birds. It has also been suggested that Dinosauria be defined as all the descendants of the most recent common ancestor of Megalosaurus and Iguanodon, because these were two of the three genera cited by Richard Owen when he recognized the Dinosauria.
Dinosaurs are divided into Ornithischia (bird-hipped) and Saurischia (lizard-hipped), depending upon the pelvic girdle structure. Ornithischian dinosaurs had a four-pronged pelvic configuration, incorporating a caudally-directed (rear-pointing) pubis bone with (most commonly) a forward-pointing process. By contrast, the pelvic structure of saurischian dinosaurs was three-pronged, and featured a pubis bone directed cranially, or forwards, only (Benton 2004b). Ornithischia includes all taxa sharing a more recent common ancestor with Triceratops than with Saurischia, while Saurischia includes those taxa sharing a more recent common ancestor with birds than with Ornithischia.

Birds are cited in the definition of Saurischia because there is an almost universal consensus among paleontologists that birds are the descendants of theropod dinosaurs (bipedal saurischian dinosaurs). Using the strict cladistical definition that all descendants of a single common ancestor are related, modern birds are dinosaurs and dinosaurs, therefore, are not extinct. Modern birds are classified by most paleontologists as belonging to the subgroup Maniraptora, which are coelurosaurs, a kind of theropods, which are saurischians, which are dinosaurs (Padian 2004).
However, referring to birds as 'avian dinosaurs' and to all other dinosaurs as 'non-avian dinosaurs' is cumbersome. Birds are still referred to as birds, at least in popular usage and among ornithologists. It is also technically correct to refer to birds as a distinct group under the older Linnaean classification system, which accepts paraphyletic taxa that exclude some descendants of a single common ancestor. Paleontologists mostly use cladistics, which classifies birds as dinosaurs, although some biologists do not.
For clarity, this article will use "dinosaur" as a synonym for "non-avian dinosaur", and "bird" as a synonym for "avian dinosaur" (meaning any animal that evolved from the common ancestor of Archaeopteryx and modern birds). The term "non-avian dinosaur" will be used for emphasis as needed.
Size

While the evidence is incomplete, it is clear that, as a group, dinosaurs were large. Even by dinosaur standards, the sauropods (a suborder or infraorder of the saurischian, "lizard-hipped," dinosaurs, including Apatosaurus and Brachiosaurus) were gigantic. For much of the dinosaur era, the smallest sauropods were larger than anything else in their habitat, and the largest were an order of magnitude more massive than anything else that has since walked the Earth. Giant prehistoric mammals such as the Indricotherium and the Columbian mammoth were dwarfed by the giant sauropods, and only a handful of modern aquatic animals approach or surpass them in size—most notably the blue whale, which reaches up to 190,000 kg (209 tons) and 33.5 m (110 ft) in length.
Most dinosaurs, however, were much smaller than the giant sauropods. Current evidence suggests that dinosaur average size varied through the Triassic, early Jurassic, late Jurassic, and Cretaceous periods (Holtz, 1998). According to paleontologist Bill Erickson, estimates of median dinosaur weight range from 500 kg to 5 metric tons; a recent study of 63 dinosaur genera yielded an average weight greater than 850 kg—comparable to the weight of a grizzly bear—and a median weight of nearly 2 tons, or about as much as a giraffe. This contrasts sharply with the size of modern mammals; on average, mammals weigh only 863 grams, or about as much as a large rodent. The smallest dinosaur was bigger than two-thirds of all current mammals; the majority of dinosaurs were bigger than all but 2% of living mammals (Erickson 2005).
Largest and smallest dinosaurs
Only a tiny percentage of animals ever fossilize, and most of these remain buried in the earth. Few of the specimens that are recovered are complete skeletons, and impressions of skin and other soft tissues are rare. Rebuilding a complete skeleton by comparing the size and morphology of bones to those of similar, better-known species is an inexact art, and reconstructing the muscles and other organs of the living animal is, at best, a process of educated guesswork. As a result, scientists will probably never be certain of the largest and smallest dinosaurs.
The tallest and heaviest dinosaur known from good skeletons is Brachiosaurus brancai (also known as Giraffatitan). Its remains were discovered in Tanzania between 1907–12. Bones from multiple similarly-sized individuals were incorporated into the skeleton now mounted and on display at the Humboldt Museum of Berlin (Colbert 1968); this mount is 12 m (38 ft) tall, 22.5 m (74 ft) long, and would have belonged to an animal that weighed between 30,000–60,000 kg (33–66 short tons, with a short ton equal to 2,000 pounds). The longest complete dinosaur is the 27 m (89 ft) long Diplodocus, which was discovered in Wyoming in the United States and displayed in Pittsburgh's Carnegie Natural History Museum in 1907.
There were larger dinosaurs, but knowledge of them is based entirely on a small number of fragmentary fossils. Most of the largest herbivorous specimens on record were all discovered in the 1970s or later, and include the massive Argentinosaurus, which may have weighed 80,000–100,000 kg (88–121 short tons); the longest, the 40 m (130 ft) long Supersaurus; and the tallest, the 18 m (60 ft) Sauroposeidon, which could have reached a sixth-floor window. The longest of them all may have been Amphicoelias fragillimus, known only from a now lost partial vertebral neural arch described in 1878. Extrapolating from the illustration of this bone, the animal may have been 58 m (190 ft) long and weighed over 120,000 kg (132 short tons) (Carpenter 2006), heavier than all known dinosaurs except possibly the poorly known Bruhathkayosaurus, which could have weighed 175,000-220,000 kg (193-243 tons). The largest known carnivorous dinosaur was Spinosaurus, reaching a length of 16-18 meters (53-60 ft), and weighing in at 9 tons (dal Sasso et al. 2006). Other large meat-eaters included Giganotosaurus, Mapusaurus, Tyrannosaurus rex and Carcharodontosaurus.
Not including modern birds like the bee hummingbird, the smallest dinosaurs known were about the size of a crow or a chicken. The theropods Microraptor and Parvicursor were both under 60 cm (2 ft) in length.
Behavior

Interpretations of dinosaur behavior are generally based on the pose of body fossils and their habitat, computer simulations of their biomechanics, and comparisons with modern animals in similar ecological niches. As such, the current understanding of dinosaur behavior relies on speculation, and will likely remain controversial for the foreseeable future. However, there is general agreement that some behaviors common in crocodiles and birds, dinosaurs' closest living relatives, were also common among dinosaurs.
The first perceived direct evidence of herding behavior was the 1878 discovery of 31 Iguanodon dinosaurs, which were thought to have perished together in Bernissart, Belgium, after they fell into a deep, flooded sinkhole and drowned (Yans et al. 2005). Despite the deposition of those skeletons being regarded now as more gradual (Van Acker 2001), other well supported mass death sites were subsequently discovered. Those, along with multiple trackways, suggest that herd or pack behavior was common in many dinosaur species. Trackways of hundreds or even thousands of herbivores indicate that duck-bills (hadrosaurids) may have moved in great herds, like the American Bison or the African Springbok. Sauropod tracks in England document that these animals traveled in groups composed of several different species (Day and Upchurch 2002). Others kept their young in the middle of the herd for defense, according to trackways at Davenport Ranch, Texas. Dinosaurs may have congregated in herds for defense, for migratory purposes, or to provide protection for their young.
Paleontologist Jack Horner's 1978 discovery of a Maiasaura ("good mother dinosaur") nesting ground in Montana demonstrated that parental care apparently continued long after birth among the ornithopods (Lessem and Glut 1993). There is also evidence that other Cretaceous period dinosaurs, like the Patagonian sauropod Saltasaurus (1997 discovery), had similar nesting behaviors, and that the animals congregated in huge nesting colonies like those of penguins. The Mongolian maniraptoran Oviraptor was discovered in a chicken-like brooding position in 1993, which may mean it was covered with an insulating layer of feathers that kept the eggs warm. Trackways have also confirmed parental behavior among sauropods and ornithopods from the Isle of Skye in northwestern Scotland (BBC 2003). Nests and eggs have been found for most major groups of dinosaurs, and it appears likely that dinosaurs communicated with their young, in a manner similar to modern birds and crocodiles.

The crests and frills of some dinosaurs, like the marginocephalians, theropods, and lambeosaurines, may have been too fragile to be used for active defense, so they were likely used for sexual or aggressive displays, though little is known about dinosaur mating and territoriality. The nature of dinosaur communication also remains enigmatic, and is an active area of research. For example, recent evidence suggests that the hollow crests of the lambeosaurines may have functioned as resonance chambers used for a wide range of vocalizations.
From a behavioral standpoint, one of the most valuable dinosaur fossils was discovered in the Gobi Desert in 1971, when paleontologists unearthed a Velociraptor attacking a Protoceratops, illustrating interspecific predation.
Paleontologists have also speculated on cannibalistic behavior, based on evidence such as toothmarks (Rogers et al. 2003).
There have not been any few indications climbing species of dinosaurs, and no indication of a burrowing species. Since the later mammalian radiation in the Cenozoic produced many burrowing and climbing species, the lack of evidence for similar dinosaur species is somewhat surprising.
Because most dinosaur species seem to have relied on land-based locomotion, a good understanding of how dinosaurs moved on the ground is key to models of dinosaur behavior; the science of biomechanics, in particular, has provided significant insight in this area. For example, studies of the forces exerted by muscles and gravity on dinosaurs' skeletal structure have offered a new understanding of how fast dinosaurs could run (Thulborn 1982), whether diplodocids could create sonic booms via whip-like tail snapping (Douglas and Young 1998), whether giant theropods had to slow down when rushing for food to avoid fatal injuries (Hecht 1998), and whether sauropods could float (Henderson 2003).
Evolution of dinosaurs
Dinosaurs appear to have diverged from their archosaur ancestors approximately 230 million years ago during the Middle to Late Triassic period, roughly 20 million years after the Permian-Triassic extinction event wiped out an estimated 95% of all life on Earth.
Radiometric dating of fossils from the early dinosaur genus Eoraptor establishes its presence in the fossil record at this time. Paleontologists believe Eoraptor resembles the common ancestor of all dinosaurs (Hayward 1997). If this is true, its traits suggest that the first dinosaurs were small, bipedal predators (Sereno et al. 1993). The discovery of primitive, dinosaur-like ornithodirans such as Marasuchus and Lagerpeton in Argentinian Middle Triassic strata supports this view; analysis of recovered fossils suggests that these animals were indeed small, bipedal predators.
The first few lines of primitive dinosaurs diversified rapidly through the rest of the Triassic period; dinosaur species quickly appeared with the the specialized features and range of sizes needed to exploit nearly every terrestrial ecological niche. During the period of dinosaur predominance, which encompassed the ensuing Jurassic and Cretaceous periods, nearly every known land animal larger than 1 meter in length was a dinosaur.
The Cretaceous-Tertiary extinction event, which occurred approximately 65 million years ago at the end of the Cretaceous period, caused the extinction of all dinosaurs except for the line that had already given rise to the first birds. Other diapsid species related to the dinosaurs also survived the event.
Study of dinosaurs
Knowledge about dinosaurs is derived from a variety of fossil and non-fossil records, including fossilized bones, feces, trackways, gastroliths (stones in the stomach), feathers, impressions of skin, internal organs, and soft tissues (Dal Sasso and Signore 1998, Schwitzer et al. 2005). Many fields of study contribute to our understanding of dinosaurs, including physics, chemistry, biology, and the earth sciences (of which paleontology is a sub-discipline).
Dinosaur remains have been found on every continent on Earth, including Antarctica. Numerous fossils of the same dinosaur species have been found on completely different continents, corroborating the generally-accepted theory that all land masses were at one time connected in a super-continent called Pangaea. Pangaea began to break apart during the Triassic period roughly 230 million years ago (Evans 1998).
History of discovery
Dinosaur fossils have been known for millennia, although their true nature was not recognized. The Chinese, whose own word for dinosaur is konglong (恐龍, or "terrible dragon"), considered them to be dragon bones and documented them as such. For example, Hua Yang Guo Zhi, a book written by Zhang Qu during the Western Jin Dynasty, reported the discovery of dragon bones at Wucheng in Sichuan Province (Zhiming 1992). In Europe, dinosaur fossils were generally believed to be the remains of giants and other creatures killed by the Great Flood.

Megalosaurus was the first dinosaur to be formally described, in 1677, when part of a bone was recovered from a limestone quarry at Cornwell near Oxford, England. This bone fragment was identified correctly as the lower extremity of the femur of an animal larger than anything living in modern times. The second dinosaur species to be identified, Iguanodon, was discovered in 1822 by the English geologist Gideon Mantell, who recognized similarities between his fossils and the bones of modern iguanas. Two years later, the Rev. William Buckland, a professor of geology at Oxford University, unearthed more fossilized bones of Megalosaurus and became the first person to describe dinosaurs in a scientific journal.
The study of these "great fossil lizards" soon became of great interest to European and American scientists, and in 1842 the English paleontologist Richard Owen coined the term "dinosaur." He recognized that the remains that had been found so far, Iguanodon, Megalosaurus and Hylaeosaurus, shared a number of distinctive features, and so decided to present them as a distinct taxonomic group. With the backing of Prince Albert of Saxe-Coburg-Gotha, the husband of Queen Victoria, Owen established the Natural History Museum in South Kensington, London, to display the national collection of dinosaur fossils and other biological and geological exhibits.
In 1858, the first known American dinosaur was discovered, in marl pits in the small town of Haddonfield, New Jersey (although fossils had been found before, their nature had not been correctly discerned). The creature was named Hadrosaurus foulkii, after the town and the discoverer, William Parker Foulke. It was an extremely important find; Hadrosaurus was the first nearly complete dinosaur skeleton found and it was clearly a bipedal creature. This was a revolutionary discovery as, until that point, most scientists had believed dinosaurs walked on four feet, like other lizards. Foulke's discoveries sparked a wave of dinosaur mania in the United States.


Dinosaur mania was exemplified by the fierce rivalry in the United States between Edward Drinker Cope and Othniel Charles Marsh, both of whom raced to be the first to find new dinosaurs in what came to be known as the Bone Wars. The feud probably originated when Marsh publicly pointed out that Cope's reconstruction of an Elasmosaurus skeleton was flawed; Cope had inadvertently placed the plesiosaur's head at what should have been the animal's tail end. The fight between the two scientists lasted for over 30 years, ending in 1897 when Cope died after spending his entire fortune on the dinosaur hunt. Marsh 'won' the contest primarily because he was better funded through a relationship with the U.S. Geological Survey (and he helped to end Cope's funding). Unfortunately, many valuable dinosaur specimens were damaged or destroyed due to the pair's rough methods; for example, their diggers often used dynamite to unearth bones (a method modern paleontologists would find appalling). The rivalry was fierce: in addition to efforts to hurt each other's reputation, there were allegations of stealing fossils, stealing workers, treaty violations, and even that Marsh dynamited a quarry rather than let Cope get the fossils.
Despite their unrefined methods, the contributions of Cope and Marsh to paleontology were vast; Marsh unearthed 86 new species of dinosaur and Cope discovered 56, for a total of 142 new species. Cope's collection is now at the American Museum of Natural History in New York, while Marsh's is on display at the Peabody Museum of Natural History at Yale University (Williams 1997).
Since 1897, the search for dinosaur fossils has extended to every continent, including Antarctica. The first Antarctic dinosaur to be discovered, the ankylosaur Antarctopelta oliveroi, was found on Ross Island in 1986, although it was 1994 before an Antarctic species, the theropod Cryolophosaurus ellioti, was formally named and described in a scientific journal.
Current dinosaur "hot spots" include southern South America (especially Argentina) and China. China in particular has produced many exceptional feathered dinosaur specimens due to the unique geology of its dinosaur beds, as well as an ancient arid climate particularly conducive to fossilization. The well-preserved feathered dinosaurs in China have further consolidated the link between dinosaurs and their conjectured living descendants, modern birds).
The current "dinosaur renaissance"

The field of dinosaur research has enjoyed a surge in activity that began in the 1970s and is ongoing. This was triggered, in part, by John Ostrom's discovery of Deinonychus, an active, vicious predator that may have been warm-blooded, and was a marked contrast to the then-prevailing image of dinosaurs as sluggish and cold-blooded. Vertebrate paleontology, arguably the primary scientific discipline involved in dinosaur research, has become a global science. Major new dinosaur discoveries have been made by paleontologists working in previously unexploited regions, including India, South America, Madagascar, and most significantly in China.
The widespread application of cladistics, which rigorously analyzes the relationships between biological organisms, has also proved tremendously useful in classifying dinosaurs. Cladistic analysis, among other modern techniques, helps to compensate for an often incomplete and fragmentary fossil record.
Classification
Dinosaurs (including birds) are archosaurs, like modern crocodilians. Archosaurs' diapsid skulls have two holes located where the jaw muscles attach, called temporal fenestrae. Most reptiles (including birds) are diapsids; mammals, with only one temporal fenestra, are called synapsids; and turtles, with no temporal fenestra, are anapsids. Anatomically, dinosaurs share many other archosaur characteristics, including teeth that grow from sockets rather than as direct extensions of the jawbones. Within the archosaur group, dinosaurs are differentiated most noticeably by their gait. Dinosaur legs extend directly beneath the body, whereas the legs of lizards and crocodylians sprawl out to either side.
Collectively, dinosaurs are usually regarded as a superorder or an unranked clade. They are divided into two orders, the Saurischia and the Ornithischia, on the basis of their hip structure. Saurischians ('lizard-hipped', from the Greek sauros (σαυρος) meaning 'lizard' and ischion (ισχιον) meaning 'hip joint') are dinosaurs that originally retained the hip structure of their ancestors. They include all the theropods (bipedal carnivores) and sauropods (long-necked herbivores). Ornithischians ('bird-hipped', from the Greek ornitheos (ορνιθειος) meaning 'of a bird' and ischion (ισχιον) meaning 'hip joint') is the other dinosaurian order, most of which were quadrupedal herbivores.
Note that the terms "lizard hip" and "bird-hip" are misnomers and misleading—birds evolved from dinosaurs with "lizard hips."
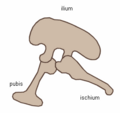
Saurischian pelvis structure (left side)

Tyrannosaurus pelvis (showing saurischian structure - left side)
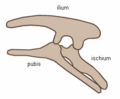
Ornithischian pelvis structure (left side).

Edmontosaurus pelvis (showing ornithischian structure - right side)




The following is a simplified classification of dinosaur families. The dagger (†) is used to indicate taxa that are extinct.
Order Saurischia

- †Infraorder Herrerasauria
- Suborder Theropoda
- †Infraorder Coelophysoidea
- †Infraorder Ceratosauria
- Family Ceratosauridae
- Family Abelisauridae
- Clade Tetanurae
- †Superfamily Megalosauroidea
- Family Megalosauridae
- Family Spinosauridae
- †Infraorder Carnosauria
- Clade Coelurosauria
- †Superfamily Tyrannosauroidea
- †Infraorder Ornithomimosauria
- †Infraorder Segnosauria
- †Infraorder Oviraptorosauria
- †Infraorder Deinonychosauria
- Family Dromaeosauridae
- Family Troodontidae
- †Superfamily Megalosauroidea

- †Suborder Sauropodomorpha
- Infraorder Prosauropoda
- Family Riojasauridae
- Family Plateosauridae
- Family Massospondylidae
- Infraorder Sauropoda
- Family Anchisauridae
- Family Melanorosauridae
- Family Blikanasauridae
- Family Vulcanodontidae
- Family Cetiosauridae
- Family Omeisauridae
- Clade Turiasauria
- Clade Neosauropoda
- Superamily Diplodocoidea
- Family Camarasauridae
- Family Brachiosauridae
- Superfamily Titanosauroidea
- Infraorder Prosauropoda
Order Ornithischia

- †Family Fabrosauridae
- †Suborder Thyreophora
- Family Scelidosauridae
- Infraorder Stegosauria
- Infraorder Ankylosauria
- Family Nodosauridae
- Family Ankylosauridae
- †Suborder Cerapoda
- Family Heterodontosauridae
- Infraorder Ornithopoda
- Family Hypsilophodontidae
- Family Iguanodontidae
- Superfamily Hadrosauroidea
- Clade Marginocephalia
- Infraorder Pachycephalosauria
- Infraorder Ceratopsia
- Family Psittacosauridae
- Family Protoceratopsidae
- Family Ceratopsidae
Areas of debate
Warm-bloodedness

A vigorous debate on the subject of temperature regulation in dinosaurs has been ongoing since the 1960s. Originally, scientists broadly disagreed as to whether dinosaurs were capable of regulating their body temperatures at all. More recently, the presence of endothermy among dinosaurs has become the consensus view, and debate has focused on the mechanisms of temperature regulation.
After dinosaurs were discovered, paleontologists first posited that they were ectothermic creatures: "terrible lizards" as their name suggests. This supposed cold-bloodedness implied that dinosaurs were relatively slow, sluggish organisms, comparable to modern reptiles, which need external sources of heat in order to regulate their body temperature. Dinosaur ectothermy remained a prevalent view until Robert T. "Bob" Bakker, an early proponent of dinosaur endothermy, published an influential paper on the topic in 1968.
Modern evidence indicates that dinosaurs thrived in cooler temperate climates, and that at least some dinosaur species must have regulated their body temperature by internal biological means (perhaps aided by the animals' bulk). Evidence of endothermism in dinosaurs includes the discovery of polar dinosaurs in Australia and Antarctica (where they would have experienced a cold, dark six-month winter), the discovery of dinosaurs whose feathers may have provided regulatory insulation, and analysis of blood-vessel structures within dinosaur bone that are typical of endotherms. Skeletal structures suggest that theropods and other dinosaurs had active lifestyles better suited to an endothermic cardiovascular system, while sauropods exhibit fewer endothermic characteristics. It is certainly possible that some dinosaurs were endothermic while others were not. Scientific debate over the specifics continues (Parsons 2001).
Complicating the debate is the fact that warm-bloodedness can emerge based on more than one mechanism. Most discussions of dinosaur endothermy tend to compare them to average birds or mammals, which expend energy to elevate body temperature above that of the environment. Small birds and mammals also possess insulation, such as fat, fur, or feathers, which slows down heat loss. However, large mammals, such as elephants, face a different problem because of their relatively small ratio of surface area to volume (Haldane's principle). This ratio compares the volume of an animal with the area of its skin: as an animal gets bigger, its surface area increases more slowly than its volume. At a certain point, the amount of heat radiated away through the skin drops below the amount of heat produced inside the body, forcing animals to use additional methods to avoid overheating. In the case of elephants, they have reduced hair, and have large ears which increase their surface area, and have behavioral adaptations as well (such as using the trunk to spray water on themselves and mud wallowing). These behaviors increase cooling through evaporation.
Large dinosaurs would presumably have had to deal with similar issues; their body size suggests they lost heat relatively slowly to the surrounding air, and so could have been what are called inertial homeotherms, animals that are warmer than their environments through sheer size rather than through special adaptations like those of birds or mammals. However, so far this theory fails to account for the vast number of dog- and goat-sized dinosaur species which made up the bulk of the ecosystems during the Mesozoic period.
Feathered dinosaurs and the bird connection
Birds and non-avian dinosaurs (dinosaurs in common, non-cladistic terminology) share many features. Birds share over a hundred distinct anatomical features with theropod dinosaurs, which are generally accepted to have been their closest ancient relatives (Mayr 2005).
Feathers

Archaeopteryx, the first good example of a "feathered dinosaur," was discovered in 1861. The initial specimen was found in the Solnhofen limestone in southern Germany, which is a lagerstätte, rare and remarkable sedimentary deposits that exhibit extraordinary fossil richness or completeness. Archaeopteryx has been considered a transitional fossil, with features intermediate between those of modern reptiles and birds. This Jurassic fossil animal is widely accepted as the earliest and most primitive known bird. However, it is so dinosaur-like that, without a clear impression of feathers in the surrounding rock, at least one specimen was mistaken for Compsognathus (Wellnhofer 1988). Fossils show the presence of wings and feathers, as in birds, but also teeth (which modern birds do not have), claws on the wings, and a long, lizard-like tail, with tail vertebrate, such as with reptiles (Mayr 2001).
The description of the first intact specimen in 1861 (two years after Charles Darwin published The Origin of Species), set off a firestorm of debate about evolution and the role of transitional fossils that endures to this day. Over the years, ten specimens of Archaeopteryx have been found, all in a limestone deposit near Solnhofen, Germany. Nonetheless, despite the significance attributed to this species, Archaeopteryx does not significantly illuminate possible transitions between reptiles/dinosaurs and birds, since it appears in the fossil record fully formed, with a lack of any transitional forms between reptiles or dinosaurs and itself.
Since the 1990s, a number of additional "feathered dinosaurs" have been found, providing even stronger evidence of the close relationship between dinosaurs and modern birds. Most of these specimens were unearthed in the Liaoning province in northeastern China, which was part of an island continent during the Cretaceous period. Though feathers have been found only in the lagerstätte of the Yixian Formation and a few other places, it is possible that non-avian dinosaurs elsewhere in the world were also feathered. There is a lack of widespread fossil evidence for feathered non-avian dinosaurs. This could be a function of the fact that delicate features like skin and feathers are not often preserved by fossilization and thus are absent from the fossil record.
The feathered dinosaurs discovered so far include Beipiaosaurus, Caudipteryx, Dilong, Microraptor, Protarchaeopteryx, Shuvuuia, Sinornithosaurus, Sinosauropteryx, and Jinfengopteryx. Dinosaur-like birds like Confuciusornis, which are anatomically closer to modern avians, have also been discovered. All of these specimens come from the same formation in northern China. The dromaeosauridae family in particular seems to have been heavily feathered, and at least one dromaeosaurid, Cryptovolans, may have been capable of flight.
Skeleton
.jpg)
Because feathers are often associated with birds, feathered dinosaurs are often touted regarding the relationship between birds and dinosaurs. However, the multiple skeletal features also shared by the two groups represent the more important link for paleontologists. Furthermore, it is increasingly clear that the relationship between birds and dinosaurs, and the evolution of flight, are more complex topics than previously realized. For example, while it was once believed that birds evolved from dinosaurs in one linear progression, some scientists, most notably Gregory S. Paul, conclude that dinosaurs such as the dromaeosaurs may have evolved from birds, losing the power of flight while keeping their feathers in a manner similar to the modern ostrich and other ratites.
Comparison of bird and dinosaur skeletons, as well as cladistic analysis, strengthens the case for the link, particularly for a branch of theropods called maniraptors. Skeletal similarities include the neck, pubis, wrist (semi-lunate carpal), arm, and pectoral girdle, shoulder blade, clavicle, and breast bone.
Reproductive biology
A discovery of features in a Tyrannosaurus rex skeleton recently provided even more evidence that dinosaurs and birds evolved from a common ancestor and, for the first time, allowed paleontologists to establish the sex of a dinosaur. When laying eggs, female birds grow a special type of bone in their limbs. This medullary bone, which is rich in calcium, forms a layer inside the hard outer bone that is used to make eggshells. The presence of endosteally-derived bone tissues lining the interior marrow cavities of portions of the Tyrannosaurus rex specimen's hind limb suggested that T. rex used similar reproductive strategies, and revealed the specimen to be female.
A dinosaur embryo was found without teeth, suggesting that some parental care was required to feed the young dinosaur. It is also possible that the adult dinosaurs regurgitated into a young dinosaur's mouth to provide sustenance, a behavior that is also characteristic of numerous modern bird species.
Lungs
Large meat-eating dinosaurs had a complex system of air sacs similar to those found in modern birds, according to an investigation which was led by paleontologist Patrick O'Connor. The lungs of theropod dinosaurs (carnivores that walked on two legs and had birdlike feet) likely pumped air into hollow sacs in their skeletons, as is the case in birds. "What was once formally considered unique to birds was present in some form in the ancestors of birds," O'Connor said (O'Connon and Claessens 2005).
Heart and sleeping posture
Modern computerized tomography (CT) scans of a dinosaur chest cavity conducted in 2000 found the apparent remnants of a complex four-chambered heart, much like those found in today's mammals and birds (Fisher et al. 2000). The idea is controversial within the scientific community, coming under-fire for bad anatomical science (Hillenius and Ruben 2004), or simply wishful thinking (Rowe et al. 2001). A recently discovered troodont fossil demonstrates that the dinosaurs slept like certain modern birds, with their heads tucked under their arms (Xu and Norell 2004). This behavior, which may have helped to keep the head warm, is also characteristic of modern birds.
Gizzard
Another piece of evidence that birds and dinosaurs are closely related is the use of gizzard stones. These stones are swallowed by animals to aid digestion and break down food and hard fibers once they enter the stomach. When found in association with fossils, gizzard stones are called gastroliths. Because a particular stone could have been swallowed at one location before being carried to another during migration, paleontologists sometimes use the stones found in dinosaur stomachs to establish possible migration routes.
Evidence for Cenozoic dinosaurs
In 2002, paleontologists Zielinski and Budahn reported the discovery of a single hadrosaur leg bone fossil in the San Juan Basin, New Mexico (United States). The formation in which the bone was discovered has been dated to the early Paleocene epoch approximately 64.5 million years ago. If the bone was not re-deposited into that stratum by weathering action, it would provide evidence that some dinosaur populations may have survived at least a half million years into the Cenozoic era) Fassett et al. 2002).
Bringing dinosaurs back to life

There has been much speculation about the use of technology to bring dinosaurs back to life. In Michael Crichton's book Jurassic Park, which popularized the idea, scientists use blood from fossilized mosquitos that have been suspended in tree sap since the Mesozoic to reconstruct the DNA of dinosaurs, filling chromosomal gaps with modern frog genes. It is probably impossible to resurrect dinosaurs in this manner. One problem with the amber extraction method is that exposure to air, water, and radiation causes DNA to decay over time, making it unlikely that such an approach would recover sufficient genetic material in an intact state.
The successful extraction of ancient DNA from dinosaur fossils has been reported on two separate occasions, but upon further inspection and peer review, neither of these reports could be confirmed (Wang et al. 1997). However, a functional visual peptide of a (theoretical) dinosaur has been inferred using analytical phylogenetic reconstruction methods on gene sequences of still-living related species (reptiles and birds) (Chang et al. 2002).
Even if dinosaur DNA could be reconstructed, it would be exceedingly difficult to "grow" dinosaurs using current technology since no closely related species exist to provide zygotes or a suitable environment for embryonic development.
Soft tissue in dinosaur fossils
One of the best examples of soft tissue impressions in a fossil dinosaur was discovered in Petraroia, Italy. The discovery was reported in 1998, and described the specimen of a small, very young coelurosaur, Scipionyx samniticus. The fossil includes portions of the intestines, colon, liver, muscles, and windpipe of this immature dinosaur (Dal Sasso and Signore 1998).
In the March 2005 issue of Science, Dr. Mary Higby Schweitzer and her team announced the discovery of flexible material resembling actual soft tissue inside a 68-million-year-old Tyrannosaurus rex leg bone from the Hell Creek Formation in Montana (Schweitzer et al. 2005). After recovery, the tissue was rehydrated by the science team.
When the fossil was treated over several weeks to remove mineral content from the fossilized bone marrow cavity (a process called demineralization), Schweitzer found evidence of intact structures such as blood vessels, bone matrix, and connective tissue (bone fibers) (Schweitzer et al. 2005). Scrutiny under the microscope further revealed that the putative dinosaur soft tissue had retained fine structures (microstructures) even at the cellular level.
The exact nature and composition of this material are not clear. Interpretation of the sample is ongoing, and the relative importance of Dr. Schweitzer's discovery is not yet clear (Schweitzer et al. 2005).
Extinction theories
The sudden mass extinction of non-avian dinosaurs, an event that occurred approximately 65 million years ago, is one of the most intriguing mysteries in paleontology. Many other groups of animals also became extinct at this time, including ammonites (nautilus-like mollusks), mosasaurs, plesiosaurs, pterosaurs, herbivorous turtles and crocodiles, most birds, and many groups of mammals.
The nature of the event(s) that caused this mass extinction has been extensively studied since the 1970s; at present, several related theories are supported by paleontologists. Though the general consensus is that an impact event was the primary cause of dinosaur extinction, some scientists cite other causes, or support the idea that a confluence of several factors was responsible for the sudden disappearance of dinosaurs from the fossil record.
Asteroid collision

The asteroid collision theory, which was first proposed by Walter Alvarez in the late 1970s, links the extinction event at the end of the Cretaceous period to a bolide (meteor) impact approximately 65.5 million years ago. Alvarez proposed that a sudden increase in iridium levels, recorded around the world in the period's rock stratum, was direct evidence of the impact. The bulk of the evidence now suggests that a 5-15 km wide bolide hit in the vicinity of the Yucatán Peninsula, creating the 170 km-wide Chicxulub Crater and triggering the mass extinction. Scientists are not certain whether dinosaurs were thriving or declining before the impact event. Some scientists propose that the meteorite caused a long and unnatural drop in Earth's atmospheric temperature, while others claim that it would have instead created an unusual heat wave.
Although the speed of extinction cannot be deduced from the fossil record alone, various models suggest that the extinction was extremely rapid. The consensus among scientists who support this theory is that the impact caused extinctions both directly (by heat from the meteorite impact) and also indirectly (via a worldwide cooling brought about when matter ejected from the impact crater reflected thermal radiation from the sun).
Multiple collisions—the Oort cloud
While similar to Alvarez's impact theory (which involved a single asteroid or comet), this theory proposes that a stream of comets was dislodged from the Oort cloud due to the gravitational disruption caused by a passing star. One or more of these objects then collided with the Earth at approximately the same time, causing the worldwide extinction. As with the impact of a single asteroid, the end result of this comet bombardment would have been a sudden drop in global temperatures, followed by a protracted cool period (Koeberl and MacLeod 2002).
Environment changes
At the peak of the dinosaur era, there were no polar ice caps, and sea levels are estimated to have been from 100 to 250 meters (330 to 820 feet) higher than they are today. The planet's temperature is also thought to have been much more uniform, with only 25 degrees Celsius separating average polar temperatures from those at the equator. On average, atmospheric temperatures were also much warmer; the poles, for example, were 50°C warmer than today (Mentzer 2018).
The atmosphere's composition during the dinosaur era was vastly different as well. Carbon dioxide levels were up to 12 times higher than today's levels, and oxygen formed 32 to 35 percent of the atmosphere, as compared to 21 percent today.
However, by the late Cretaceous, the environment was changing dramatically. Volcanic activity was decreasing, which led to a cooling trend as levels of atmospheric carbon dioxide dropped. Oxygen levels in the atmosphere also started to fluctuate and would ultimately drop considerably. Some scientists hypothesize that climate change, combined with lower oxygen levels, might have led directly to the demise of many species. If the dinosaurs had respiratory systems similar to those commonly found in modern birds, it may have been particularly difficult for them to cope with reduced respiratory efficiency, given the enormous oxygen demands of their very large bodies.
In popular culture
By human standards, dinosaurs were creatures of fantastic appearance and often enormous size. As such, they have captured people's imaginations and become an enduring part of human popular culture. Dinosaur exhibitions and displays at parks and museum exhibits around the world both cater to, and reinforce, this public interest. The popular preoccupation with dinosaurs is also reflected in a broad array of fictional and non-fictional works.
Notable examples of older fictional works featuring dinosaurs include Arthur Conan Doyle's book The Lost World; the 1933 film King Kong; and Godzilla.
ReferencesISBN links support NWE through referral fees
- BBC. 2003. Dinosaur family footprints found BBC News. December 2, 2003. Retrieved July 18, 2019.
- Benton, M. J. 2004a. Origin and Relationships of Dinosaurs. In D. B. Weishampel, P. Dodson, and H. Osmólska, eds., The Dinosauria, 2nd ed., 7-19. Berkeley: University of California Press. ISBN 0520067266
- Benton, M. J. 2004b. Vertebrate Paleontology. London: Blackwell Publishers. ISBN 978-0632056378
- Carpenter, K. 2006. Biggest of the big: A Critical Re-evaluation of the Mega-sauropod Amphicoelias fragillimus. In J. R. Foster, and S. G. Lucas, eds., Paleontology and Geology of the Upper Jurassic Morrison Formation. New Mexico Museum of Natural History and Science Bulletin 36: 131-138.
- Chang, B. S. W., K. Jönsson, M. A. Kazmi, M. J. Donoghue, and T. P. Sakmar. 2002. Recreating a Functional Ancestral Archosaur Visual Pigment. Molecular Biology and Evolution 19: 1483-1489.
- Colbert, E. H. 1968. Men and Dinosaurs: The Search in Field and Laboratory. New York: E. P. Dutton & Company. ISBN 0140212884
- Day, J. J., and P. Upchurch. 2002. Sauropod Trackways, Evolution, and Behavior. Science 296: 1659.
- dal Sasso, C., S. Maganuco, E. Buffetaut, and M. A. Mendez. 2006. New Information on the Skull of the Enigmatic Theropod Spinosaurus, With Remarks on its Sizes and Affinities. Journal of Vertebrate Paleontology 25(4): 888-896.
- dal Sasso, C., and M. Signore. 1998. Exceptional Soft-tissue Preservation in a Theropod Dinosaur from Italy. Nature 292: 383-387.
- Douglas, K., and S. Young. 1998. The Dinosaur detectives. New Scientist 2130: 24.
- Erickson, W. C. 2005. On the Origin of Dinosaurs and Mammals. Retrieved July 18, 2019.
- Evans, J. 1998. Ultimate Visual Dictionary - 1998 Edition. Dorling Kindersley Books. 66-69. ISBN 1871854008
- Farlow, J. O., and M. K. Brett-Surman. 1997. Preface. In J. O. Farlow and M. K. Brett-Surman, eds., The Complete Dinosaur. Indiana University Press: Bloomington and Indianapolis. ISBN 0253333490
- Fassett, J., R. A. Zielinski, and J. R. Budahn. 2002. Dinosaurs That Did Not Die: Evidence for Paleocene Dinosaurs in the Ojo Alamo Sandstone, San Juan Basin, New Mexico. In C. Koeberl and K. MacLeod, eds., Catastrophic Events and Mass Extinctions: Impacts and Beyond. Special Paper - Geological Society of America 356: 307-336.
- Fisher, P. E., D. A. Russell, M. K. Stoskopf, R. E. Barrick, M. Hammer, and A. A. Kuzmitz. 2000. Cardiovascular Evidence for an Intermediate or Higher Metabolic Rate in an Ornithischian Dinosaur. Science 288: 503-505.
- Fountain, H. 2006. Many More Dinosaurs Still to be Found. New York Times September 12, 2006.
- Gunter, V. A. 2001. Bernissart iguanodons and their significance. Cleveland Museum of Natural History. Retrieved July 18, 2019.
- Hayward, T. 1997. The First Dinosaurs. Dinosaur Cards. Orbis Publishing Ltd.
- Hecht, J. 1998. The Deadly Dinos That Took a Dive. New Scientist 2130.
- Henderson, D. M. 2003. Effects of Stomach Stones on the Buoyancy and Equilibrium of a Floating Crocodilian: A Computational Analysis. Canadian Journal of Zoology 81: 1346-1357.
- Hillenius, W. J., and J. A. Ruben. 2004. The Evolution of Endothermy in Terrestrial Vertebrates: Who? When? Why? Physiological and Biochemical Zoology 77: 1019-1042.
- Holtz, T. R. 2000. Classification and Evolution of the Dinosaur Groups. In G. S. Paul, ed., The Scientific American Book of Dinosaurs. New York: St. Martin's Press, 140-168. ISBN 0312262264
- Koeberl, C., and K. G. MacLeod. 2002. Catastrophic Events and Mass Extinctions. Geological Society of America. ISBN 0813723566
- Langer, M. C., F. Abdala, M. Richter, M. J. Benton. 1999. A Sauropodomorph Dinosaur from the Upper Triassic (Carnian) of Southern Brazil. Comptes Rendus de l'Academie des Sciences, Paris: Sciences de la terre et des planètes 329: 511-517.
- Lessem, D., and D. F. Glut. 1993. The Dinosaur Society's Dinosaur Encyclopedia. Random House Inc. ISBN 0679417702
- Mayr, E. 2001. What Evolution Is. New York: Basic Books. ISBN 0465044255
- Mayr, G., B. Pohl, and D. S. Peters. 2005. A Well-Preserved Archaeopteryx Specimen with Theropod Features. Science 310: 1483-1486.
- Mentzer, A.P. 2018. The Climate in the Jurassic Era Sciencing. Retrieved July 18, 2019.
- O'Connor, P. M., and L. P. A. M. Claessens. 2005. Basic Avian Pulmonary Design and Flow-through Ventilation in Non-avian Theropod Dinosaurs. Nature 436: 253.
- Owen, R. 1842. Report on British Fossil Reptiles. Part II. Plymouth, England: Report of the British Association for the Advancement of Science.
- Padian, K. 2004. Basal Avialae. In D. B. Weishampel, P. Dodson, and H. Osmólska, eds., The Dinosauria, second edition. Berkeley: University of California Press. 210-231. ISBN 0520242092
- Padian, K., and Philip J. Currie. 1997. Encyclopedia of Dinosaurs. Academic Press. ISBN 0122268105
- Parsons, K. M. 2001. Drawing Out Leviathan. Indiana University Press. ISBN 0253339375
- Paul, G. S. 2000. The Scientific American Book of Dinosaurs. St. Martin's Press. ISBN 0312262264
- Paul, Gregory S. 2002. Dinosaurs of the Air: The Evolution and Loss of flight in Dinosaurs and Birds. Baltimore: The Johns Hopkins University Press. ISBN 0801867630
- Rogers, R. R., D. W. Krause, and K. C. Rogers. 2003. Cannibalism in the Madagascan Dinosaur Majungatholus atopus. Nature 422: 515-518. See commentary on the article. Retrieved July 18, 2019.
- Rowe, T., E. F. McBride, P. C. Sereno, D. A. Russell, P. E. Fisher, R. E. Barrick, and M. K. Stoskopf. 2001. Dinosaur with a heart of stone. Science 291: 783.
- Schweitzer, M. H., J. L. Wittmeyer, and J. R. Horner. 2005. Soft-tissue Vessels and Cellular Preservation in Tyrannosaurus rex. Science 307: 1952-1955.
- Sereno, P. C., C. A. Forster, R. R. Rogers, and A. M. Monetta. 1993. Primitive Dinosaur Skeleton from Argentina and the Early Evolution of Dinosauria. Nature 361: 64-66.
- Strauss, B. 2018. Learn About the Different Dinosaur Periods ThoughtCo. Retrieved July 18, 2019.
- Thulborn, R.A. 1982. Speeds and gaits of dinosaurs Palaeogeography, Palaeoclimatology, Palaeoecology 38(3–4) (July 1982): 227-256. Retrieved July 18, 2019.
- Wang, S. C., and P. Dodson. 2006. Estimating the diversity of dinosaurs. Proceedings of the National Academy of Sciences USA 103(37): 13601-13605. Retrieved July 18, 2019.
- Wang, H., Z. Yan, and D. Jin. 1997. Reanalysis of Published DNA Sequence Amplified from Cretaceous Dinosaur Egg Fossil. Molecular Biology and Evolution 14: 589-591.
- Weishampel, D. B. 2004. The Dinosauria, second edition. University of California Press. ISBN 0520242092
- Wellnhofer, P. 1988. Ein neuer Exemplar von Archaeopteryx. Archaeopteryx 6: 1-30.
- Williams, P. 1997. The Battle of the Bones. Dinosaur Cards. Orbis Publishing Ltd.
- Xu, X., and M. A. Norell. 2004. A New Troodontid Dinosaur from China With Avian-like Sleeping Posture. Nature 431: 838-841.
- Yans, J., J. Dejax, D. Pons, C. Dupuis, and P. Taquet. 2005. Palaeontological and Geodynamical Implications of the Palynological Dating of the Wealden Facies Sediments of Bernissart (Mons Basin, Belgium). C. R. Palevol 4: 135–150.
- Zhiming, D. 1992. Dinosaurian Faunas of China. Beijing: China Ocean Press. ISBN 3540520848
External links
All links retrieved January 29, 2024.
- New Dinosaurs A site dedicated to providing images and facts about dinosaurs and other prehistoric life.
- Dinosaurs & other extinct creatures From the Natural History Museum. Popular site, well illustrated dino directory.
- Mesozoic Marine Monsters of the Mangahouanga The Discoveries of Dr. Joan Wiffen, New Zealand's Dinosaur Lady.
- Dinosaurs: Facts and Fiction From the United States Geological Survey. Popular overview.
- Dinosaur From the BBC.
- Planet Dinosaur From the BBC.
- The Dinosauria From UC Berkeley Museum of Paleontology Detailed information.
- Dinosaurs: News & Pictures of Dinosaur Types All about Dinosaurs with current featured articles.
- Putting Dinosaurs' Noses Back Where They Really Were.
- Prehistoric Planet From PaleoClones. Current dino news.
- DinoBuzz Are birds really Dinosaurs?
Credits
New World Encyclopedia writers and editors rewrote and completed the Wikipedia article in accordance with New World Encyclopedia standards. This article abides by terms of the Creative Commons CC-by-sa 3.0 License (CC-by-sa), which may be used and disseminated with proper attribution. Credit is due under the terms of this license that can reference both the New World Encyclopedia contributors and the selfless volunteer contributors of the Wikimedia Foundation. To cite this article click here for a list of acceptable citing formats.The history of earlier contributions by wikipedians is accessible to researchers here:
The history of this article since it was imported to New World Encyclopedia:
Note: Some restrictions may apply to use of individual images which are separately licensed.